


Le sue dimensioni molto grandi vengono interpretate come risposta all’assenza di predatori terrestri ed una protezione da quelle aeree. La presenza di una manopola carpale nel carpo metacarpo prossimale indica anche un comportamento combattivo per questa grande specie di uccelli terrestri. Qui sotto una traduzione parziale dello studio in inglese pubblicato su http://rsos.royalsocietypublishing.org.
1. Introduzione
Dal momento che le opere di Darwin e Wallace, lo studio delle isole è stato un potente strumento in molti studi evolutivi, e gli uccelli sono un gruppo chiave per comprendere i cambiamenti biologici in ambienti insulari. Gli uccelli sulle isole spesso mostrano aumenti di dimensioni, variazioni allometriche e, in alcuni casi, inadattabilità al volo. Quest’ultimo aspetto è conosciuto in diversi gruppi di uccelli, come gli ibis, i cormorani, le rotaie, i piccioni, i pappagalli, i passeriformi e gli uccelli acquatici [ 1 – 4 ].
Gli uccelli acquatici del gruppo degli Anseriformi, comprende Anhimidae (Sud America) Screamers, Anseranatinae (Australasian gazza-oche) e Anatidi con una distribuzione in tutto il mondo [ 5 ]. I fossili degli anatidi sono molto ricchi e comprendono diverse specie endemiche, che si sono evolute senza la possibilità di volare in condizioni insulari, sia sulle isole oceaniche remote [ 4 , 6 – 8 ] così come in isole intermedie come quelle del Mediterraneo [ 9 – 11 ]. La maggior parte degli anatidi insulari mostrano il grado lieve di riduzione delle ali verso impossibilità al volo [ 7 , 12 – 14 ], ma in alcuni casi, risultavano estremamente predisposti ad uno stile di vita terrestre con un grande aumento delle dimensioni del corpo, come i moa-nalos delle Hawaii (Chelychelynechen e Thambetochen) [ 4 ] e della Nuova Zelanda (Cnemiornis) [ 15 ]. Tra le specie insulari giganti di Anseriformi vi era il Garganornis ballmanni, una specie recentemente descritta grazie ai depositi risalenti al Neogene presenti sul Gargano (Italia meridionale) sulla base di un unico tiobiotarso distale [ 16 ].

E’ quindi possibile descrivere nuovi elementi scheletrici di G. ballmanni da Gargano a Scontrone, sud e centro Italia, rispettivamente ( figura 1 ), le principali località della cosiddetta “Paleobioprovincia Puglia-Abruzzo” [ 17 ], che durante il Miocene è stata caratterizzata da condizioni insulari e un’alta concentrazione di fauna endemica. Questi nuovi reperti consentono di chiarire gli adattamenti peculiari di Garganornis ad uno stile di vita terrestre e confermare la sua impossibilità al volo,già in precedenza ipotizzata [ 16 ]. Inoltre, G. ballmanni rappresenta il primo uccello fossile trovato a Scontrone, e un nuovo taxon terrestre condiviso tra Scontrone al Gargano, dimostrando che era diffusa nella Palaeobioprovincia Puglia-Abruzzo.
2. Materiali e metodi
2.1. aree di studio
L’area del Gargano (Foggia, Italia meridionale) durante il tardo Miocene era un dominio insulare che faceva parte della Puglia-Abruzzo Palaeobioprovincia [ 17 ]. Nel sedimento riempimento della rete carsica, esposta nelle cave sfruttando il Calcare di Bari risalente al Mesozoico ritrovato nei pressi di Apricena, una grande quantità di vertebrati resti è stata trovata, a dimostrazione della grande diversità della fauna che abitano questo dominio insulare [ 18 ]. La frazione fossile a Scontrone si trova sul Monte Civita, vicino al villaggio di Scontrone (L’Aquila, Italia centrale). I resti fossili sono stati trovati nella parte inferiore del Calcarenite di Scontrone: carbonati marini marginali che rappresentano la fase iniziale della trasgressione tortoniana, in un intervallo rappresentato da banchi di sabbia costiere [ 19 ]. Gargano e Scontrone rappresentano le più importanti località fossili di questo palaeobioprovincia [ 17 ], che è stato caratterizzata da associazioni vertebrati molto particolari con macro altamente endemici e micromammiferi, rettili e uccelli [ 16 – 18 , 20 – 25 ], tutti raggruppate nel cosiddetto gruppo della fauna Mikrotia [ 18 , 26 ]. Vale la pena ricordare che l’associazione di vertebrati fossili di questa palaeobioprovincia è anche caratterizzato dall’assenza di predatori terrestri, con l’eccezione del riccio molto particolare deinogalerix [ 25 ], la lontra Paralutra [ 27 ] e un coccodrillo [ 28 ], quest’ultimo noto attraverso pochissimi resti fossili.
Nonostante i vari taxa comuni di mammiferi e rettili condivisi fra Gargano e Scontrone, nemmeno un taxon del ricco assemblaggio di uccelli del Gargano è stato trovato a Scontrone o in altre località della palaeobioprovincia fino ad oggi [ 17 ]. Ciò è probabilmente dovuto alle caratteristiche tafonomiche dei depositi Scontrone [ 29 ] e ai sedimenti molto duri che incorporano le ossa, che rendono difficile trovare e raccogliere le ossa più piccole e più fragili durante gli scavi.
2.2. metodi
Il materiale fossile viene memorizzato nel Dipartimento di Scienze della Terra dell’Università degli Studi di Firenze, Firenze, Italia (DSTF), il Museo di Geologia e Paleontologia dell’Università degli Studi di Torino (MGPT-PU), il Naturmuseum di Augusta, Augusta, Germania (NMA ), la Naturalis Biodiversity center, Leiden, Paesi Bassi (RGM) e il Centro di Documentazione Paleontologico hoplitomeryx, Scontrone, Italia (SCT). Gli esemplari fossili sono stati confrontati con gli scheletri di animali moderni memorizzati nel Laboratoire de Géologie de Lyon, Université Claude-Bernard Lyon 1, Lione, Francia (FSL), il Dipartimento di Scienze della Terra dell’Università degli Studi di Torino, Torino, Italia (MPOC ), il Museo di Storia naturale, Londra e Tring, Regno Unito (NHMUK) e nel Museo nazionale di Storia naturale Ditsong, Pretoria, Sud Africa (TM). La terminologia osteologici segue Baumel & Witmer [ 30 ]. La circonferenza del tibiotarso dell’Anseriforme fossile presso il Museo Nazionale di Storia Naturale di Washington DC, Stati Uniti d’America, sono stati misurati avvolgendo un sottile cinturino di carta intorno alla parte più sottile del tibiotarso e misurando la circonferenza minima con pinze scorrevoli calibrate con l’approssimazione di 0,05 mm. Massa corporea è stata poi stimata utilizzando i minimi quadrati stime di regressione di Iwaniuk et al . [ 31 ].
3. paleontologia sistematica
Anseriformes Wagler 1831
Anatidi Vigors 1825
Garganornis ballmanni Meijer 2014
Le ossa di Garganornis ballmanni Meijer 2014 dal tardo Miocene del Gargano ( A-F ) e Scontrone ( g ), Italia. ( Un ) carpometacarpus destro DSTF-GA 49, vista ventrale; ( B ) ha lasciato carpometacarpus NMA 504/1801, vista ventrale; ( C ) prossimale pedale falange RGM 261.535, vista dorsale; ( D ) prossimale pedale falange RGM 261945, vista dorsale; ( E ) mesiale pedale falange MGPT-PU 135.536, vista dorsale; ( F ) tarsometatarsus sinistra RGM 425.554, vista dorsale; ( G ) a destra tarsometatarsus SCT 23, vista dorsale. Barra di scala rappresenta 1 cm.
Figura 3:
Disegni delle ossa di Garganornis ballmanni Meijer, 2014; tutte le ossa sono dal tardo Miocene del Gargano tranne uno ( k – m ), che è dal tardo Miocene di Scontrone, Italia. ( A-C ) carpometacarpus destro DSTF-GA 49 in ventrale ( una ), dorsale ( b ) e (prossimale c ) vista; ( D , e ) carpometacarpus sinistra NMA 504/1801 in ventrale ( d ) e dorsali ( e vista); ( F , g ) ricostruzione grafica della carpometacarpus di Garganornis ballmanni riferisce campioni DSTF-GA 49 e NMA 504/1801 in ventrale ( f dorsale () e g visualizzazioni); ( H-j ) destro tibiotarsus DSTF-GA 77 in craniale ( h ), caudale ( i ) e prossimale ( j ) vista; ( K-m ) un tarsometatarsus SCT 23 a dorsale ( k ), plantare ( l ) e (laterali m visualizzazioni); ( N, o , s, t ) sinistra tarsometatarsus RGM 425.554 a dorsale ( n ), plantare ( o ), distale ( s ) e mediale ( t visualizzazioni); ( P , u ) pedale prossimale falange prossimale RGM 261.945 a ( p ) e (dorsale u ) vista; ( Q , v , w ) prossimale pedale falange RGM 261.535 a prossimale ( q ), dorsale ( v ) e plantare ( w ) vista; ( R , x , y ) mesiale pedale falange MGPT-PU 135.536 a prossimale ( R ), dorsale ( x ) e plantari ( Y visualizzazioni). Barra di scala rappresenta 3 cm. I disegni realizzati da Ursula B. Göhlich.
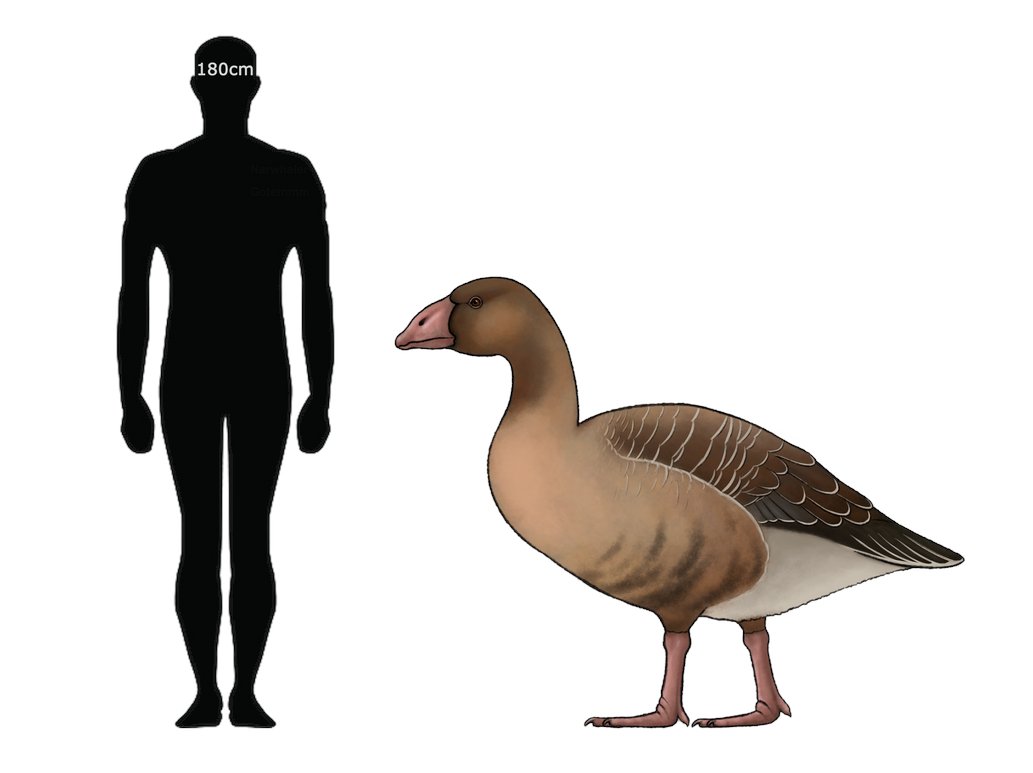
Ricostruzione artistica di Garganornis ballmanni Meijer 2014 sulla base dei resti fossili descritti di recente. Questa ricostruzione illustra una generica Oca Paleoartica occidentale con brevi e robusti tarso-metatarsali, le dita dei piedi corte e ali molto brevi in base agli elementi noti di Garganornis ballmanni. Illustrazione a cura di Stefano Maugeri.
Olotipo: distale tibiotarso sinistro RGM 443.307 dal tardo Miocene del Gargano (Posticchia 5), l’Italia meridionale.
Nuovo materiale da Gargano: destro carpometacarpus DSTF-GA 49 (F15), estremità prossimale; carpometacarpus sinistra NMA 504/1801 (Fina 2), privo di estremità prossimale; destra tibiotarsus DSTF-GA 77 (F15), danneggiato estremità prossimale; tarso-metatarsali sinistra RGM 425.554 (San Giovannino), estremità distale; trochlea metatarsi III di un tarso-metatarsali destra RGM 425.943 (Chirò 14A); indeterminati falangi pedale MGPT-PU 135.356 (fessura senza nome), RGM 261.535 (Pizzicoli 12), RGM 261945 (fessura senza nome).
Nuovo materiale da Scontrone: tarsometatarsus destra, SCT 23, quasi completa con estremità prossimale e distale danneggiate e manca troclea metatarsi IV.
Misure: DSTF-GA 49: larghezza prossimale, 27,7 mm; profondità prossimale, 10,5 mm. NMA 504/1801: lunghezza totale come conservata, 53,6 millimetri. DSTF-GA 77: larghezza prossimale, 43,2 millimetri; profondità prossimale, 31,0 millimetri (stimato). SCT 23: lunghezza totale di esemplari conservati, 168,2 millimetri; larghezza distale, il 22,5 mm (stima); larghezza del pozzo a metà lunghezza, 18,3 millimetri. RGM 425.554: larghezza distale, 47,5 millimetri; profondità distale, 24,8 millimetri (stimato). MGPT-PU 135.356: lunghezza totale, 29,0 millimetri; larghezza prossimale, il 13,4 mm profondità prossimale, 11,6 millimetri; larghezza distale, 9.1 mm profondità distale, 7,7 millimetri; larghezza del pozzo a metà lunghezza, 9.9 mm. RGM 261.535: lunghezza totale, 55,1 millimetri; larghezza prossimale, 19,6 millimetri; profondità prossimale, 17,7 millimetri; larghezza distale, 13,3 millimetri; profondità distale, 11,9 millimetri; larghezza del pozzo a metà lunghezza, 11.3 mm. RGM 261945: larghezza prossimale, 20,9 millimetri; profondità prossimale, 16,2 millimetri; larghezza del pozzo a metà lunghezza, 9.7 mm.
Distribuzione cronologica: G. ballmanni è ora riportato dalla località tortoniana di Scontrone, in un’età di circa 9 Ma [ 19 ], per le otturazioni tardo Miocene scissura di Gargano, per i quali di età compresa tra 6 e 5,5 Ma è stata recentemente proposta [ 32 ]. Attraverso il sistema di rete carsico del Gargano, i resti di ballmanni G. ora sono stati trovati in diversi ripieni si sviluppa su varie fasi cronologiche delineati da Masini et al . [ 18 , 23 ] dalla fase 1b al 3c Phase. In particolare, l’attribuzione cronologica alla fase 2b del pedale falange di recente raccolto MGPT-PU 135.356 è stato fatto dopo che lo studio di un associati Mikrotia esemplare, un roditore di fondamentale importanza per valutare le relative età dei campioni dal Gargano fessura-riempimento sistema [ 18 ].
Osteology comparativa: Il prossimale carpometacarpus DSTF-GA 49 (figure 2 a e 3 a-c ) è molto particolare, con i due lati della troclea carpalis notevolmente estese lateralmente, rendendo la parte prossimale dell’osso molto piatto rispetto ad altre forme di anseriformi. La presenza di un processus extensorius parallelo alla troclea prossimale e non inclinato sul ventre, un relativamente ampio e non indicato pisiformis processus, e una piccola dorsale manopola per la fovea carpalis caudalis permette la sua attribuzione al gruppo degli Anatidi. Sulla parte superiore della extensorius processus della carpometacarpus prossimale DSTF-GA 49, si può osservare una piccola escrescenza ossea, simile per forma e dimensioni a quella osservata in altri Anseriformes, nel femminili Pezophaps solitaria, e in Goura cristata (Columbidae), ma meno sviluppato rispetto a quelli del maschio di P. solitaria , e sicuramente non è uno sperone come in Anhimidae, Plectropterus (Anatidi) e Vanellus (Charadriidae) [ 33 ]. Campione NMA 504/1081 (figure 2 B e 3 d-e ) manca l’estremità prossimale, ad eccezione delle parti distali degli carpalis trochlea e la alularis processus quadrato. L’albero è molto ridotto in lunghezza con il spatium intermetacarpale molto breve, come dimostrato dalla distalis la sinostosi parzialmente conservata del metacarpale. Nonostante l’estremità distale dell’osso risulti essere incompleto, questa è comunque ben conservata e la sinostosi metacarpalie è ben sviluppata, il che indica che la lunghezza totale dell’osso non era molto più lunga nel campione conservato. La presenza di una fossa particolare sul lato ventrale del carpometacarpus laterali al pisiformis processus, che si riscontra anche in altri Anatidi anche se meno evidente, e altri dettagli morfologici rendono possibile ricostruire graficamente l’osso completo ( figura 3 f-g ) e per confermare la brevità relativa del carpometacarpo. Questo è il primo osso dell’ala noto per G. ballmanni .
Il prossimale tibiotarso DSTF-GA 77 ( figura 3 h-j ) è altamente danneggiato, ma mostra tre ben distinte retropatellaris Fosse e un flexoria fossa profonda, che sono compatibili con la morfologia deglii anatidi. Il olotipo tibiotarsus RGM 443.307, già descritto in dettaglio da Meijer [ 16 ], mostra chiaramente le sue affinità con gli Anatidi, piuttosto che con altri Anseriformi.
Il tarsometarso SCT 23 (figura 2 g e 3 k-m ), anche se mal conservato, è morfologicamente simile al più grande degli Anatidi, ma sempre più grande del tutt’oggi esistente Cygnus Cygnus e il fossile Cy. falconeri , il taxon estinto gigante vissuto tra Sicilia e Malta [ 34 ]. In particolare, il tarsometaro è breve e robusto con un hypotarsus centrale, dedotta dalla parte distale del hypotarsus che si conserva, l’assente solco flexorius, un extensorius solco ben marcata, e una vasta estremità distale con la arrotondata troclea metatarsi III. L’estremità prossimale inclusa la hypotarsus non viene mantenuto. Tuttavia, come la tuberosità tibiale craniale musculi è visibile sul campione, la maggiore lunghezza dell’osso completo può essere prevista per non essere molto maggiore della lunghezza totale del campione. Il plantare crista lateralis è ben sviluppato, più sporgenti plantarly che in Cygnus e Anser , ma simile a Cereopsis ; il solco di extensorius è molto profondo, più profondo che in Cygnus e nelle altre grandi Anseriformi, e si estende distalmente al punto centrale dell’osso. La parte distale della diafisi delle due esemplari fossili è piatta appena prossimale al trochleae, che non è comune nei Anseriformi, ma simile a quella osservata in Cereopsis . Il metatarsi tre trochleae, conservato nel RGM 425.554 (figure 2 F e 3 n-q ), sono latero-mediale allineati in vista distale con la troclea metatarsi II e non inclinato mediale, come nella maggior parte dei Anseriformi. Inoltre, il trochleae metatarsi II e IV sono più uguali in estensione distale rispetto alla maggior parte delle Anseriformi, ma simile a quanto osservato nel Anhimidae in Cereopsis , e in alcuni esemplari inediti della estinta Cy. falconeri conservato nel NHMUK (MP 2014, osservazione personale). E ‘importante notare che la disposizione della troclea distale differisce da quella in altri taxa grande corpo come gru (Gruidae) e cicogne (Ciconiidae). La morfologia generale dell’estremità distale del SCT 23 e RGM 425.554 sono molto bene una confrontabili tra loro sia nella forma del trochleae conservati e nella zona pianeggiante prossimalmente alla trochleae. L’avifauna del Gargano non riporta altri uccelli di queste dimensioni, e pertanto è ipotizzabile che entrambi i campioni possano rappresentare la stessa specie.
Le falangi pedale (figure 2 c-e e 3 r-y ) sono simili a quelli della più grande Anatidi [ 15 , 34 ], ma più robusto senza forti impressioni legamento.
Osservazioni: La composizione faunistica della Palaeobioprovincia di Puglia-Abruzzo è stata ampiamente studiata [ 16 , 18 , 20 – 22 , 24 ]. La più grande taxa aviaria mostra la presenza di specie Accipitridae freudenthali Garganoaetus e Tytonidae Tyto gigantea [ 20 , 21 ]. Nonostante le loro dimensioni, le ossa sono più piccole di quelle qui descritte. Inoltre, come parte di una revisione di tutta l’associazione degli uccelli provenienti dalla Palaeobioprovincia, in atto da uno di noi (MP), tutti i campioni sono stati controllati e tutti i resti degli anatidi sono stati pubblicati [ 24 ]. Nessuno di loro è paragonabile per dimensioni con G. ballmanni .
La morfologia delle ossa qui descritte, anche se fortemente modificata, si adatta meglio con quello degli Anatidi e la loro dimensione è ben compatibile con G. ballmanni . Nonostante le differenti località, si suppone appartengano tutti a G. ballmanni .
Lo studio del materiale recentemente scoperto permette di chiarire la morfologia complessiva di G. ballmanni e suoi adeguamenti ad uno stile di vita terrestre. La morfologia del tarso-metatarso è diverso dalla maggior parte dei Anseriformi esistenti e fossili per quanto riguarda il solco extensorius proporzionalmente più profondo e distale più sviluppato, il dorso-plantarly appiattito parte distale del pozzo, e metatarsi II trochlea distale e latero-mediale quasi allineati e IV. Queste caratteristiche richiamano la morfologia tarsometatarsus di Anhimidae e Cereopsis , tra i taxa non volanti che più si sono adattati alla terraferma all’interno della corona-gruppo degli Anseriformi. Inoltre, le falangi pedale conservati sono forti e brevi rispetto a quelli delle oche e cigni nuotatori.
Così, le caratteristiche morfologiche di G. ballmanni rivelano il suo adattamento a un terrestre, non acquatico, stile di vita e suggeriscono che probabilmente questi viveva in ambienti aperti o aridi, come regolarmente osservato in altra isola fortemente modificata dagli Anseriformi [ 4 , 15 ].
4. Discussione
4.1. affinità tassonomiche
L’analisi del tibiotarso olotipo di G. ballmanni ha rivelato le sue dimensioni molto grandi, molto più grandi di Cy. olor . Alcuni personaggi del tibiotarsus sono anche visto in linee estinte di Paleogene e Neogene uccelli, come dromornithidae, gastornithidae e Presbyornithidae. In particolare, G. ballmanni condivide con gastornithidae un tibiotarsus con le intercondylaris fosse larghe e l’apertura circolare del lato distale del extensorius Canalis, ma si differenzia per la mancanza di un forte solco m. fibularis e per l’estremità distale del canalis non extensorius posizionati più medialmente come in gastornithidae. Tuttavia, la presenza di questi vecchi taxa nel Neogene del sud Italia è improbabile, in quanto non vi è alcuna prova di sopravvivenza del Paleogene elementi faunistici nella Paleobioprovincia di Puglia-Abruzzo. Alternativamente, e più probabile che le morfologie peculiari mostrate dall’olotipo di G. ballmanni siano il risultato di una evoluzione convergente di questa specie in un ambiente insulare, dove carnivori mammiferi erano estremamente rari se non assente [ 25 , 27 ]. Questa teoria è supportata anche dalla morfologia delle ossa appena descritte, che sono molto più simili agli Anatidi rispetto ad gastornithidae e altri staminali gruppi di Anseriformes. E ‘anche interessante notare che carpometacarpo di G. ballmanni mostra una certa somiglianza con quella di Gastornis sp. da Louvois (Francia), mentre è molto diverso da quello da Cernay (Francia) [ 35 ]. Come per l’olotipo di tibiotarso [ 16 ], questa somiglianza non riflette alcuna relazione filogeneticae tra Garganornis e Gastornis, ma piuttosto indica evoluzione convergente. La morfologia dei nuovi resti fossili, anche se fortemente modificata e non completamente conservata, combinata con quella del olotipo di G. ballmanni , conferma la sua attribuzione alla famiglia degli Anatidi, l’unica corona-gruppo anseriformi registrato nel Neogene europeo.
Le forti modificazioni morfologiche di G. ballmanni e il piccolo numero e la cattiva conservazione dei resti fossili non consentono una ipotesi per quanto riguarda i suoi più vicini parenti non endemici continentali tra gli Anatidi e le affinità filogenetiche di questo taxon endemica.
4.2. Impossibilità al volo e stima della massa corporea
Nella descrizione originale di G. ballmanni , Meijer [ 16 , p. 22] ipotizzò che era impossibile volare sulla base della sue grandi dimensioni, rispetto agli Anatidi volanti. Il materiale appena descritto, in particolare carpometacarpo modificato, conferma questa ipotesi e ci permette di ricostruire l’aspetto di vita di G. ballmanni ( figura 4 ). Infatti, la lunghezza stimata della carpometacarpo di G. ballmanni è proporzionalmente molto più breve di quello delle grandi specie esistenti volanti di Anseriformes, e troppo ridotta per consentire alla specie di volare ( figura 3 f-g ). Inoltre, l’appiattimento dell’estremità prossimale e la proiezione laterale dei carpalis troclea sono caratteristiche uniche di G. ballmanni possibilmente correlate ad una modifica del carpometacarpo seguito della perdita della sua capacità di volo. Infatti, la carpale trochleae articolare le carpometacarpus con l’ulna e ossa carpali guida l’estensione e flessione delle ali. La loro debolezza in forma indicherebbe meno movimento dell’articolazione del polso, come osservato nella carpometacarpus di altre specie di volanti [ 15 , 31 , 36 ]. Nella maggior parte degli Anseriformi volanti, le importanti modifiche dell’ala si verificano tra le ali, radio e ulna, piuttosto che nel carpometacarpo [ 11 ]. La forte riduzione del carpometacarpus, che è comune ai volanti Rallidae e Columbidi [ 2 , 4 , 37 ], è stata registrato solo nelle specie altamente modificate chendytes lawi [ 12 ], Cnemiornis [ 15 ], Thambetochen [ 4 ] e nel recentemente descritto Shiriyanetta hasegawai [ 7 ].
L’impossibilità al volo è più frequentemente interpretata come una risposta adattativa per l’assenza o la diversità dei predatori terrestri. Tuttavia, nel caso di G. ballmanni, questo taxon era ancora esposto ai predatori aerei, che erano molto abbondanti nella zona durante il Miocene [ 20 , 21 ]. La presenza di grandi predatori volanti, come ad esempio l’aquila freudenthali Garganoaetus ed il barbagianni T. gigantea , può essere una delle ragioni principali dietro il grande aumento delle dimensioni del corpo di G. ballmanni, insieme con la tendenza generale di insulare grandi dimensioni Anatidi di diventare erbivori terrestri. Ciò è in contrasto con insulare flightless piccole dimensioni Anatidae che rimangono acquatiche e che mostrano una riduzione delle dimensioni del corpo [ 6 , 12 ]. Sulla base della circonferenza minima della tibiotarsus holotype (60,2 millimetri), Meijer [ 16 , p. 22] viene stimata la massa corporea di G. ballmanni a 22,3 kg, più grande di qualsiasi anseriforme vivente. Va notato che la circonferenza minima dell’olotipo non è la circonferenza minima dell’osso stesso, come la circonferenza minima si trova più prossimali sull’albero. Regolazione per questo, Meijer [ 16 ] invece ha dato una stima gamma di 15.3-22.3 kg, in cui G. ballmanni è ancora più pesante di anatidi esistenti. Infatti, tutte le misure delle ossa di G. ballmanni che potremmo paragonare a recenti e fossili grandi dimensioni di Anatidi hanno rivelato la sua dimensione molto grande, indicando che la massa del corpo reale è probabilmente molto più vicina al valore massimo suggerito da Meijer [ 16 , p. 22]. Purtroppo, gli elementi appena descritti non consentono una nuova stima, ovvero di come tarsometatarso è coinvolta in diverse funzioni oltre alla locomozione, come l’alimentazione e posatoi, e quindi non è un buon indicatore di massa corporea [ 31 ], e gli elementi a battente non sono mai considerati in questa analisi, in particolare, quelli di uccelli non volatori.
Le grandi dimensioni G. ballmanni risultano particolarmente evidenti in confronto con altri wildfowls insulari, come è stimato essere uno dei più grandi Anseriformi mai esistito ( tabella 1 ). In particolare, all’interno della regione del Mediterraneo, l’estinto Maltese Swan Cy. falconeri raggiungeva grandi dimensioni, e la sua dimensione è stimata da 15,8 a 16,38 kg (media di 16,09 kg) di Northcote [ 34 ]. Le isole Hawaii erano un tempo la casa del grande moa-nalo, ma le loro dimensioni corporee stimati sono nettamente inferiore a quella di G. ballmanni ( tabella 1 ). Le stime per la Nuova Zelanda Oche Cnemiornis vanno da 8.03 a 15.55 kg (in media, 12,18 kg) per Cn. gracile, e da 14,5 a 20,36 kg (media di 17,46 kg) per Cn. calcitrans [ 8 ].
Tabella 1.
Stime di massa corporea di Garganornis ballmanni Meijer 2014 in confronto con altri Anseriformi insulari estinti.
4.3. adattamento di combattimento
Oltre alla sua lunghezza ridotta, l’estremità prossimale del carpometacapo di G. ballmanni mostra evidenti modificazioni morfologiche del extensorius processus. Sul lato esterno della extensorius processus della carpometacarpo prossimale DSTF-GA 49, una piccola manopola carpale è presente ( figura 2 a ). E ‘di piccole dimensioni e non vi è alcuna prova che avrebbe potuto diventare uno sperone, come in Anhimidae, ma è simile a quelli osservati in Cygnus , Anser e altri Anatidi [ 38 ], tra cui Cy. falconeri (MP 2014, osservazione personale sulla base di materiale non descritta in NHMUK). Gli speroni carpali o manopole, quando presenti, sono quasi esclusivamente situati sul extensorius processus e sono sempre utilizzati come armi [ 33 ]. Molti Anseriformi presentano speroni sulle ali o sulle manopole in relazione a comportamenti di lotta, come ad esempio Anhimidae, Tachyeres , Chloephaga , Alopochen , Cereopsis e altri Anatidi [ 38 ]. La presenza di questa manopola in G. ballmanni dovrebbe pertanto essere correlata con comportamento di lotta alare.
5. conclusione
I dati qui presentati confermano la validità tassonomica di G. ballmanni come una specie fortemente modificata di corona-gruppo Anatidi endemica al tardo Miocene nella Paleobioprovincia di Puglia-Abruzzo del centro-sud Italia, sulla base dei resti fossili rinvenuti nella zona del Gargano e a Scontrone.
Il nuovo materiale descritto, anche se non perfettamente conservato, fornisce nuovi dettagli osteologici della morfologia di G. ballmanni , che conferma la sua impossibilità al volo e al suo estremo adattamento ad un terrestre, non acquatico, stile di vita. Inoltre, abbiamo osservato evidenze di comportamento di ali usate per combattere.
Garganornis ballmanni dimostra anche il forte carattere endemico del tardo Miocene nella Puglia-Abruzzo in Palaeobioprovincia, già suggerito dai mammiferi e uccelli [altamente modificati 17 ]. Il Mar Mediterraneo si conferma così il suo ruolo di centro di speciazione e le sue isole possono essere quindi essere state un tipo intermedio di isola, con la composizione faunistica a metà strada tra quelle oceaniche e continentali, caratterizzato dalla presenza da un povero numero di mammiferi con alto grado di endemismo e senza carnivori terrestri, comportando una diversificazione dell’avifauna con taxa altamente modificati [ 11 ].
(Fonte: http://rsos.royalsocietypublishing.org/content/4/1/160722)
Qui sotto un servizio televisivo che affronta l’argomento:
Segui tutti gli aggiornamenti, clicca “MI PIACE” sulla Pagina di Paleo Puglia Reporter: